准噶尔麻蜥的繁殖方式:先有蛋还是先有蜥蜴?

先有鸡,还是先有蛋?这是一个妇孺皆知的古老谜题,迄今似乎仍无明确的答案,仁者见仁,智者见智。如果从动物繁殖方式演化的角度,那么这个问题并不难回答。几乎可以肯定,卵生出现在胎生之前,也就是先有“蛋”后有“鸡”。
今天我们从准噶尔麻蜥的繁殖方式说起,来聊聊类似的一个话题,先有蜥蜴还是先有蛋?(Which came first: the lizard or the egg?)
如先前科考日志《走过荒漠——追寻准噶尔麻蜥的前世今生》所记,准噶尔麻蜥(图1、图2、图3)不仅是额尔齐斯河流域荒漠戈壁的蜥蜴代表,也堪称该流域的特有爬行动物。细心的读者可能注意到了“密点麻蜥”与“密点麻蜥复合体”这两个术语,难免要问:它们有什么隐秘的共性呢?准噶尔麻蜥和它们到底有什么关联?简单地说,它们的突出的共性在于特别的繁殖模式⸺不是下蛋(卵生),而是产仔(胎生)。

图1 准噶尔麻蜥 (雄性,阿勒泰市巴里巴盖乡,2022.7.19,郭宪光拍摄)

图2准噶尔麻蜥 (雌性,产后体虚消瘦,阿勒泰市红墩镇汗德尕特乡,2022.7.18,郭宪光拍摄)

图3 准噶尔麻蜥(幼体,阿勒泰市红墩镇汗德尕特乡,2022.7.18,郭宪光拍摄)
为了弄清准噶尔麻蜥的身世,笔者曾于2018年蒙古科学考察期间,专程前往乌兰巴托拜访了霍尔洛·孟和巴雅尔Khorloogiin Munkhbayar(1940-),他是该物种的发现和命名者之一。
我们知道,卵生(Oviparity,egg-laying)和胎生(Viviparity, live bearing)是脊椎动物两种最基本的繁殖方式。要么母体产下卵,让后代在孵出之前在卵内继续生长;要么新个体一直待在母体内,直到长成一个更完整的幼体。当然,二者最大区别是母体在胚胎发育过程中能量投入的差异。对卵生物种而言,母体产卵,随后卵在外界环境中孵化至卵内胚胎完全发育而孵出幼体;而胎生繁殖模式则是母体将受精卵滞留在子宫或输卵管内直至胚胎发育完全,由母体直接分娩产出。
读到这里,可能有人不禁纳闷:卵胎生又是啥情况?
卵胎生(Ovoviviparity)是过时的概念,历史上用来描述“介于卵生和胎生之间”的繁殖模式。有鳞目爬行动物(蜥蜴和蛇)中约20%的已知物种可以“卵胎生”。近年来发现,传统以为“卵胎生不会为胚胎提供气体交换以外的物质支持”的原因是实验精度太差。没有胎盘的“卵胎生”同样能提供气体交换以外的物质支持。因此,“卵胎生”应该属于胎生。
根据胚胎发育的营养来源,生育方式可以分为卵性营养(Lecithotrophy)和胎性营养(Matrotrophy)两种类型。卵性营养物种胚胎发育的营养来自卵黄,而胎性营养物种胚胎发育的营养由母体通过胎盘供应。卵生物种都属于卵性营养,而胎生物种可据胚胎发育的营养来源划分为卵性营养胎生(即传统称为“卵胎生”)和胎性营养胎生,不同胎生物种处于卵性−胎性营养连续谱的不同位点上。绝大多数爬行动物处于极端卵性营养一端,但极少数胎生石龙子(如眼点石龙子或称铜蜥Chalcides ocellatus)胚胎发育显著依赖于胎性营养。
麻蜥属(Eremias)已报道42个物种,其中大多数物种的繁殖方式是卵生。最早被发现的繁殖方式为胎生的是密点麻蜥E. multiocellata。密点麻蜥是由著名的动物学家阿尔伯特·甘瑟Albert Günther(图5)于1872年,根据蒙古南部戈壁省的一号标本(Desert of Gobi, on the route from Sume to the Tola [= Tuul] river)所描述。后来,更多的胎生物种(亚种)被陆续报道。由于这些营胎生繁殖方式的麻蜥之间的进化关系扑朔迷离,剪不断、理还乱,学界为交流需要,便以“密点麻蜥复合体”(Eremias multiocellata complex)来统称它们(Eremchenko et al., 1992; Eremchenko & Panfilov, 1999; Guo et al., 2011)。
密点麻蜥复合体广布于我国西北、蒙古、俄罗斯图瓦、吉尔吉斯斯坦、哈萨克斯坦东部等地区。长期以来,由于该复合体具有复杂的种内形态变异和种间相似性,物种界定困难,历经了漫长的分类学修订,有多达18个种/亚种被提出。
笔者在系统梳理国内外文献基础上,识别了该属的8个胎生物种,即密点麻蜥、莎车麻蜥E. yarkandensis Blanford,1875、荒漠漠蜥E. przewalskii(Strauch, 1876) 、四额鳞麻蜥E. quadrifrons(Strauch, 1876)、喀什麻蜥E. buechneri Bedriaga,1907、天山麻蜥E. stummeri Wettstein,1940、纳伦麻蜥E. szcezerbaki Eremchenko & Panfilov,1992和科克沙尔麻蜥E. kokshaalensis Eremchenko & Panfilov,1999。部分胎生物种如下所示(图4)。

图4 麻蜥属部分胎生物种. A, 天山麻蜥; B, 喀什麻蜥; C, 科克沙尔麻蜥; D, 莎车麻蜥; E, 纳伦麻蜥; F, 密点麻蜥; G, 荒漠麻蜥. (A-D, 刘金龙拍摄;E, 引自Sindaco & Eremchenko, 2008; F, G, 郭宪光拍摄)
2017年,密点麻蜥复合体中的一个隐存新种⸺准噶尔麻蜥从所谓的密点麻蜥中分出。与其近缘种相比,准噶尔麻蜥具有明显的遗传和形态分化。基于线粒体COI基因序列,准噶尔麻蜥与同属其他物种的遗传距离为3.2%(与喀什麻蜥)到11.1%(与甘肃山丹的密点麻蜥)。
在蒙古准噶尔戈壁,准噶尔麻蜥栖息在海拔2400-2600m,喜欢岩石半沙漠或山地沙漠栖息地,偶尔有稀疏的植被:梭梭(藜科)、锦鸡儿(豆科)、白刺(白刺科)、红砂(柽柳科)和芨芨草(禾本科)(图5)。密点麻蜥更喜欢低海拔的沙漠栖息地,山脚下有松散的沙子和沙丘,而准噶尔麻蜥栖息在丘陵地区、岩石斜坡、露头或峡谷。当温度升至18°C以上时(通常是4月底),麻蜥会在冬眠后复出,并且通常活跃到10月初。准噶尔麻蜥大约4月底到5月交配、育种,7 月中旬出生1-4个蜥仔。在生育之前,有学者观察到怀孕的雌蜥会迁移到具有较密集的蒿属(菊科)植被的较低沙地,这可能为新生儿提供了更好的栖息地,在那里它们与密点麻蜥分享相同的生境。

图5 准噶尔麻蜥模式产地。(A)岩石露头,准噶尔麻蜥的典型栖息地 (B)密点麻蜥的典型栖息地。
曾经有学者认为,麻蜥的胎生繁殖方式可能不止一次起源(Shine, 1985),但近年的一项研究明确了麻蜥属中的胎生物种形成一个进化枝(Guo et al., 2011),这支持了麻蜥属中的胎生类群形成单系,为一次起源。此观点也得到了后来增加取样/扩充数据后的研究所支持(Orlova et al., 2017; Liu et al., 2021; Tian & Guo, 2022)。而麻蜥属中的胎生类群形成的进化枝是嵌在卵生种之间,也就是说麻蜥属中的卵生种并未形成单系群(图6)。可见,从繁殖方式的演化角度看,麻蜥属的卵生起源时间在胎生之前;先有卵生,后有胎生。这样一来,可以认为麻蜥属的演化历史中,先有蛋后有蜥蜴。
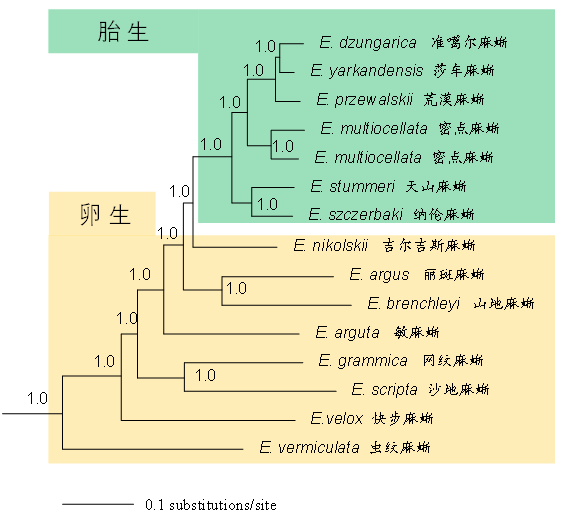
图6 基于线粒体基因组的麻蜥属部分物种的贝叶斯系统发育树(改自Tian & Guo, 2022)
那么,从麻蜥属繁殖方式的演化趋势可否推广到整个有鳞类爬行动物(包括蜥蜴、蛇和蚓蜥)吗?
有鳞类约有20%的胎生物种,胎生繁殖方式独立起源了约115次。Wright et al. (2015)通过稳健的祖先性状重建,推导出有鳞类的共同祖先是卵生的,随后向胎生的进化转变很常见,但向卵生的逆转却很罕见(图7)。具有最强系统发育支持的三个假定的卵生逆转发生在阿拉伯沙蚺 Eryx jayakari 和巨蝮属蛇Lachesis(分布于中美洲及南美洲森林地区的蝮蛇),还有滑喉蜥Liolaemus calchaqui。可见,从繁殖方式的演化角度看,的确是先有蛋后有蜥蜴。

图7 有鳞类(蜥蜴和蛇)繁殖模式的祖先状态重建,分支颜色从蓝色(支持卵生)到红色(支持胎生) (引自Wright et al., 2015)。
为什么一些蜥蜴会进化出胎生的繁殖方式呢?
爬行动物中的有鳞类的胎生起源通常被认为与寒冷气候下的繁殖方式特化有关。换言之,冷气候假说(cold-climate hypothesis)认为,胎生是动物为适应寒冷气候而演化出来的特殊繁殖模式,这是因为胎生方式能显著改善胚胎发育环境,从而有效降低了外界不良环境对胚胎发育的影响,因而能够大大提高后代的存活率和适合度。当然,胎生繁殖模式同样存在劣势,比如胚胎发育影响母体的摄食和运动能力,增加母体死亡率,降低母体繁殖输出能力。Guo et al. (2011) 使用分子钟技术来估算麻蜥不同进化枝的分歧时间,并提出其胎生支系(即密点麻蜥复合体)的起源演化(约500万年前)可能与青藏高原隆升及其引起的环境气候变化密切相关。这种观点也得到了我们后续研究的进一步支持(李大江,2015;刘金龙,2019)。
另一方面,在与爬行动物胎生进化的选择压力相关的假说中,预见性假说(Tinkle & Gibbons, 1977)和母体操纵假说(Shine, 1985)也备受关注。预见性假说(predictability hypothesis)提出,寒冷或温带地区的环境变化提供了导致卵子保留和胎生进化的选择性压力。冷气候假说和预见性假说均无法回答为什么在温暖或可预测的气候中,胎生的繁殖策略也在爬行动物中如此成功的问题。
母体操纵假说(maternal manipulation hypothesis),则认为孕期母体能通过行为调温,辅以生理调温使得自身体温发生调整,这样能为胚胎发育提供相对较高且稳定的适宜热环境;而且母体通过体温调整诱导的后代表型特征的变异能增强后代的适合度;该假说被认为能适用于任何情况或任何一个物种,近年来越来越受到研究者们的普遍关注和验证。
Li et al. (2009) 以荒漠麻蜥为材料,验证了母体操纵假说的主要预测,即雌性应在妊娠期间改变选定的体温,从而为胚胎发育提供最佳热条件,并且由母体体温调节确定的表型特征应增强后代的适应性。Li et al. (2009) 的研究首次证明母体操纵假说适用于寒冷气候的胎生爬行动物。
然而,最近的一项研究,支持了胎生的进化起源与古气候和谱系多样化一致(Recknagel et al., 2021)。他们发现,稳定和持久的寒冷气候条件与有鳞类动物由卵生向胎生的转变有关,这种繁殖模式的性状进化不能与有鳞类的谱系多样化分开。
说到这里,可能有人要问:是否有既能下蛋(卵生),又能产仔(胎生),也就是“卵生和胎生并存”的蜥蜴和蛇?答案是有,这些称为双模繁殖或双生殖(bimodal reproduction)种类。
大千世界,无奇不有!当然,双模繁殖是很罕见的:大约11,349种现生的有鳞类动物(截至2022年3月),仅有9个被可靠地描述为双模繁殖,(图8)。包括一些蜥蜴:黑尾条唇石龙子Glaphyromorphus nigricaudis , 东南石龙子Lerista bougainvillii,红尾石龙子Madascincus igneocaudatus ,三趾石龙子Saiphos equalis,海角石龙子Trachylepis capensis及胎蜥Zootoca vivipara ,和蛇:锯鳞蝰蛇Echis carinatus,棕带水蛇Helicops angulatus 及灰腹草蛇Psammophylax variabilis。尽管 Blackburn (2015) 认为这些物种被“可靠地”描述为双模繁殖方式,但由于相对缺乏研究,其中一些物种的分类可能并不健全。例如锯鳞蝰蛇E. carinatus在不同的环境中分布广泛,包括热带雨林、沙漠和高山地区。因此,这样的双模“物种”可能实际上是物种复合体,具有尚未确定的跨越地理边界的隐存异域物种形成。除了那些有强有力证据的物种,人们需要进一步的研究来确认这些物种的双模繁殖。

图8 九个可双模繁殖的蜥蜴和蛇。从A−I分别为黑尾条唇石龙子Glaphyromorphus nigricaudis,东南石龙子Lerista bougainvillii,红尾石龙子Madascincus igneocaudatus,三趾石龙子Saiphos equalis,海角石龙子Trachylepis capensis,胎蜥Zootoca vivipara,锯鳞蝰蛇Echis carinatus,棕带水蛇Helicops angulatus和灰腹草蛇Psammophylax variabilis。除图片F由郭宪光拍摄,其余均来自网络。
胎蜥Z. vivipara、东南石龙子 L. bougainvilii、三趾石龙子S. equalis 和棕带水蛇H. angulatus 的双模繁殖证据最强,有数篇到许多同行评审的出版物记录了它们的双模繁殖证据,而上面列出的其他物种的证据仅限于一些难以验证的报告,这些报告依赖于不太详细的系统发育信息。在得到证实的情况下,种内的双模繁殖为研究繁殖策略的演化提供了独特的机会,因为我们可以直接比较种内的卵生和胎生个体,而不必考虑物种形成所引起的混杂影响。这些双生殖类群为了解卵生和胎生之间主要进化转变提供了绝佳的素材,而这些转变是动物进化历史上的一项根本性创新。
19. Wright AM, et al. 2015. Which came first: the lizard or the egg? Robustness in phylogenetic reconstruction of ancestral states. J. Exp. Zool. (Mol. Dev. Evol.), 324B: 504-516.
来源:新疆三调额尔齐斯河流域生态系统与生物多样性调查项目



















 鲁公网安备37020202370214号
鲁公网安备37020202370214号